流行病学
流行病学
流行病学:
1.传染源与宿主 动物莱姆病是一种人畜共患病,现已查明有30多种野生动物(鼠、鹿、兔、狐、狼等),49种鸟类及多种家畜(狗、牛、马等)可作为本病的动物宿主。其中啮齿类动物由于其数量多、分布广及感染率高是本病的传染源。美国以野鼠中的白足鼠和驯养动物中的白尾鹿为主,我国报告有黑线姬鼠、黄胸鼠、褐家鼠、白足鼠等。因此认为本病疫源地的存在是伯氏疏螺旋体通过动物-蜱-动物的传播循环而建立起来的,然而,我国从小型啮齿动物胎鼠分离到病原体,表明垂直传播也是疫源地维持的重要方式之一。鸟类有很大散布蜱类的作用,特别是一些候鸟。近来调查我国黑龙江与四川省某些地区大型动物,发现狗的感染率为38%~60%、牛为18%~32%、羊为17%~61%之间,这些大动物宿主感染后,血清中可查到抗体,但不表现症状,可能在维持媒介的种群数量上起着重要作用。患者仅在感染早期血液中存在病原体,因而作为本病传染源的意义不大,但含有螺旋体的血液,经常规处理及血库4℃贮存48天仍有感染性,应警惕经输血传播的可能。
2.传播媒介 本病传播媒介为节肢动物-硬蜱,欧洲疫源地主要为蓖籽硬蜱(lxodes ricinus),北美东部疫源地主要为肩突硬蜱(I.scapularis,过去误定为达敏硬蜱I.dammini),北美西部疫源地主要为太平洋硬蜱(I.pacificus)。东亚疫源地在我国东北林区全沟硬蜱(I.persulcatus)为优势蜱种(占80%),带螺旋体率高达20%~50%,因而成为北方林区主要传播媒介。长江中、下游林区疫源地二棘血蜱(Haemaphysalis bispinosa)为优势蜱种(占85%),带螺旋体率为16%~40%,提示为该地区的重要生物媒介。我国南方疫源地的粒形硬蜱(I.granulatus)、寄麝硬蜱(I.moschiferi)均分离到莱姆病螺旋体,也是本病传播媒介。
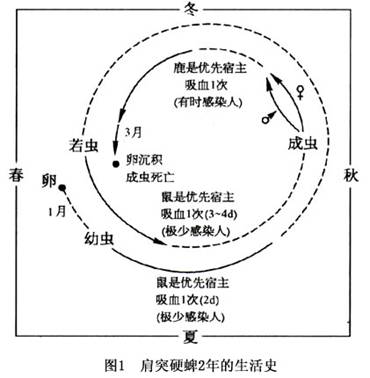
硬蜱的生活史包括三方面,即未成熟期(蚴虫、若虫)、成虫期及寄生宿主类型。观察肩突硬蜱2年的生活史,需经鼠、鹿等宿主才完成。幼虫(6腿)阶段极少使人感染,主要使人感染的是若虫(8腿)至成虫阶段(图1)。调查证明若虫是莱姆病的主要传播虫期,因其季节消长与莱姆病流行一致,且有14%~17%的病原感染率,成虫虽然感染率可高达30%~100%,但活动期在冬季,也易被人发现,故是次要传播虫期。蜱也可经卵传播螺旋体。在一般情况下,硬蜱常在森林附近或沿着动物足迹或小道,爬到植物(草或灌丛)尖端,一旦宿主接触到这些植物,蜱就能爬入宿主叮咬吸血。由于蜱细小,叮咬又无痛,常未被注意。
3.人群易感性 人群对本病原体普遍易感。重疫区可有10%~20%的居民受过感染。人体感染后可表现临床上的莱姆病或无症状的隐性感染,两者比例大致为1∶1。感染后可出现高效价的特异性IgM和IgG抗体,前者多在4~6个月内降至正常水平;后者可保持数月甚至数年。也有报道在血清特异性IgG抗体升高3~4年后仍出现莱姆病临床症状,因而此IgG抗体对人体是保护作用还是致病作用,尚待研究。
4.流行特征
(1)地区分布:莱姆病几乎在世界各地都存在,特别是在北半球分布广泛。全球已有50多个国家报道均有本病发生,其中以美国最多,病例已遍及49个州。据美国疾病控制中心从1982年开始监测以来,至今已有10万病例报道。欧洲各国每年也有5万例以上患者,在日本、埃及、南非等国也有病例报道。我国于1985年在黑龙江省海林县首次发现本病患者,1988年从患者血液中分离到病原体以来,至今已有27个省(市、区)报道发现本病,18个省(市、区)存在莱姆病自然疫源地。东北林区、西北林区、内蒙古林区为主要流行区。流行地区发病率每年为20~100/10万人,在大兴安岭、小兴安岭、长白山、天山、阿尔泰山等林区人群感染率为10%以上,在秦岭以南林区为5%~10%,在平原地区为5%以下。
(2)时间分布:莱姆病的发病时间有一定的季节性,每年有两个感染高峰期,即6月与10月,其中以6月份最明显。但在近太平洋地区,多数患者发病时间为1~5月间。我国东北林区为4~8月份,福建林区为5~9月份。其季节性发病高峰与当地蜱类的数量及活动高峰相一致。
(3)人群分布:发病以青壮年居多,与职业相关密切。以野外工作者、林业工人感染率较高。据报道疫区室外工作人员劳动一天后有40%被蜱叮咬史,或可从其皮肤、衣服等处找到蜱。室外消遣活动如狩猎、垂钓和旅游等均可增加感染本病的危险性。
病因
病因:莱姆病的病原体在1982年由Burgdorferi和Barbour等首先证实是一种新种疏螺旋体,称为伯氏包柔螺旋体(B.Burgdorferi),简称伯氏疏螺旋体。伯氏疏螺旋体是一种单细胞疏松盘绕的左旋螺旋体,长10~40μm,宽0.2~0.3μm,有3~7个疏松和不规则的螺旋,两端稍尖,是包柔螺旋体属中菌体最长而直径最窄的一种。运动活泼,可有扭转、翻滚、抖动等多种方式。革兰染色阴性,吉姆萨或瑞氏染色呈淡红的蓝色,Eosin Thiazin染色呈青紫色到浅紫色,镀银染色能使螺旋体着色良好。电镜下可见外膜和鞭毛(7~12根不等),鞭毛位于外膜与原生质之间,故又称内鞭毛(endoflagellum),与运动有关。在微需氧条件下,30~34℃在BSK-Ⅱ(Barbour Stoenner Killy-Ⅱ)培养基中生长良好,生长缓慢,一般需2~5周才可在暗视野显微镜下查到。该螺旋体有鞭毛与外膜两种抗原性蛋白:①鞭毛蛋白(flagellin),分子量为41×103,编码基因于染色体上,具有很强的抗原性和免疫原性,最早致机体产生特异性IgM抗体,其高峰滴度常在感染后6~8周,以后下降;②外膜由脂蛋白微粒组成,具有抗原性的外膜表面蛋白有OspA(outer surface protein A,分子量为31×103~32×103),OspB(分子量为34×103~36×103)及OspC(体外培养不表达,分子量为21×103),可使机体产生特异性IgG和Iga抗体,从感染后2~3个月开始,滴度逐渐增加,并可保持多年。外膜蛋白在疾病过程中可发生抗原性变异。
长期来认为莱姆病的病原体只有伯氏疏螺旋体一个种,近来依据5S-23SrRNA基因间隔区MseI限制性片段,结合DNA-DNA杂交同源性分析了世界各地分离的莱姆病菌株,至少有10个基因种(genospecies),其中可以引起莱姆病的至少有三个基因种:①狭义伯氏疏螺旋体(B.burgdorferi sensu stricto),以美国、欧洲为主;②伽氏疏螺旋体(B.garinii),以欧洲和日本为主;③阿弗西尼疏螺旋体(B.afzelii),亦从欧洲和日本分离出。我国分离的大部分菌株的蛋白图谱更接近于欧洲菌株,以伽氏和阿弗西尼疏螺旋体占优势。
莱姆病螺旋体在潮湿及低温情况下抵抗力较强,但对热、干燥和一般消毒剂均能使其灭活。
发病机制
发病机制:伯氏疏螺旋体主要存在于蜱的中肠憩室部位,当蜱叮咬人时,可从涎腺内或中肠所含螺旋体通过反流至吸食腔,然后侵入人体皮肤的微血管,经血流至全身各器官组织。然而该病原体引发
菌血症期较短,血液中螺旋体量也不多,但可引起如此多器官及多系统的损害,其致病机制可能是多因素综合的结果。1998年已发现该螺旋体有两种黏附素(adhesion),即DbpA(decorin binding protein A)和DbpB,通过黏附素使螺旋体结合到皮肤和其他器官组织细胞的胶原蛋白相关的细胞外基质蛋白多糖上,使细胞发生病变。伯氏疏螺旋体细胞壁中有脂多糖(LPS)组分,具有类似内毒素的生物学活性;及其外膜表面蛋白Osp A,Osp B,Osp C具有重要的致病力和侵袭力。螺旋体又可诱导宿主细胞释放细胞因子,这些细胞因子可以加重病变组织的炎症。
螺旋体进入皮肤约数日后,即引起第一期的局部皮肤原发性损害,受损皮肤的浅层及深层血管周围有浆细胞和淋巴细胞浸润,表现为慢性游走性红斑(ECM),螺旋体的LPS成分会使患者出现全身症状及肝
脾肿大等。ECM组织切片上可见上皮增厚,轻度角化伴单核细胞浸润,表皮层水肿,无化脓性及肉芽肿性反应。当螺旋体经血循环感染各组织器官后,进入第二期(播散病变期),以中枢神经系统(特别为脑神经)和心脏受损为主的病变。在大脑皮质血管周围及脑神经尤其面神经、动眼神经及展神经,心脏组织中有单核细胞浸润等。发病持续数月以上,则进入第三期(持续感染期),以关节、皮肤病变及晚期神经损害为主。可见关节呈增生性侵蚀性滑膜炎,伴血管增生,滑膜绒毛肥大,纤维蛋白沉着,单核细胞浸润。骨与软骨也有不同程度的侵蚀性破坏。皮肤萎缩、脱色或出现胶原纤维组织束增粗,排列紧密,类似硬皮病损害及萎缩性肢皮炎。神经系统主要为进行性脑脊髓炎和轴索性脱髓鞘病变,血管周围有淋巴细胞浸润,血管壁增厚,胶原纤维增生。